In vitro plant regeneration is the process of growing plants using tissue culture techniques. Plant regeneration can be achieved by culturing explants to form organs (organogenesis), or by developing plant embryos (somatic embryogenesis). Both techniques can lead to regenerating entire plants.
This article summarizes most of the important aspects of plant regeneration using somatic embryogenesis and organogenesis. In this article, you will find information about the differences, types, advantages, and disadvantages between somatic embryogenesis and organogenesis. I also describe the factors influencing each process and some biotech applications for both techniques.
What is Organogenesis and Somatic Embryogenesis?
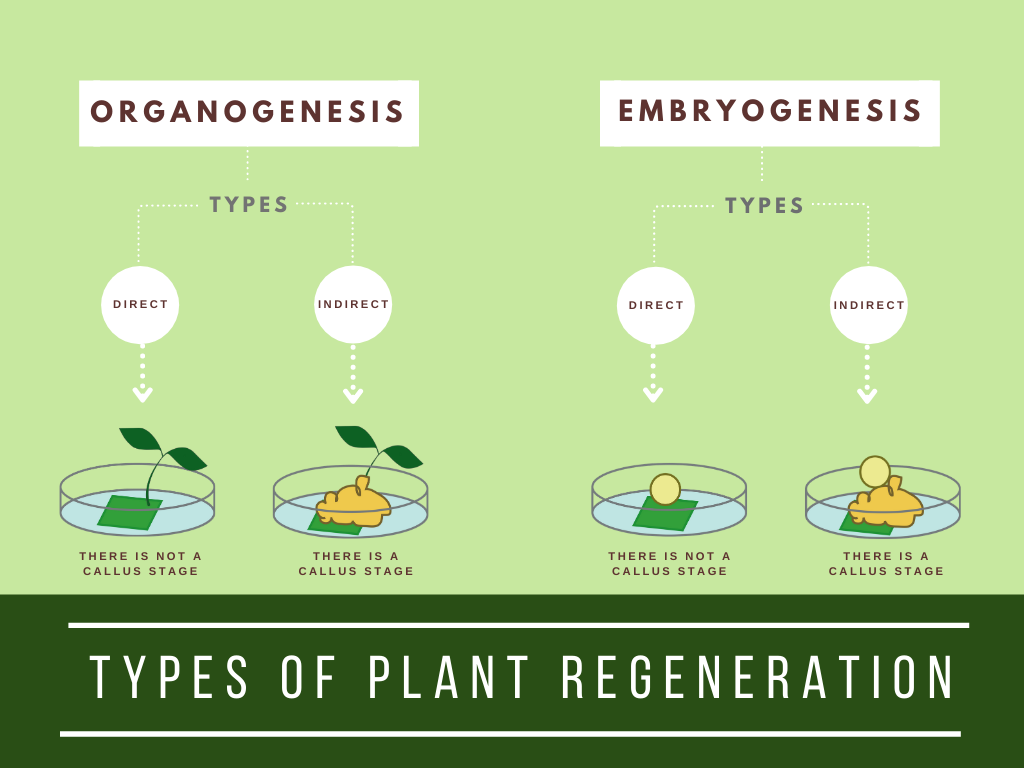
In plant tissue culture, organogenesis is the formation of organs from the cultured explants (plant material such as roots, leaves and flowers).The organogenesis process is where the plant organs, either shoots or roots, are developed.
Embryogenesis is the process of forming and developing embryos (Bhatia and Bera, 2015). Plant embryos are the area of the seed where the plant’s roots, stem and leaves start their earliest formation.
The in vitro and in vivo plant embryogenic processes are very different from each other. In nature, a mother plant is crossed with a parent plant to produce a seed; this is sexual hybridization.
In tissue culture, sexual hybridation does not occur. Instead bipolar structures that are embryo-like can regenerate from somatic cells (any plant cell other than a gamete or germ cell) (Arnold et al., 2002). Because of the embryo origin in tissue culture, this process is named somatic embryogenesis (SE).
Types of Organogenesis and somatic embryogenesis
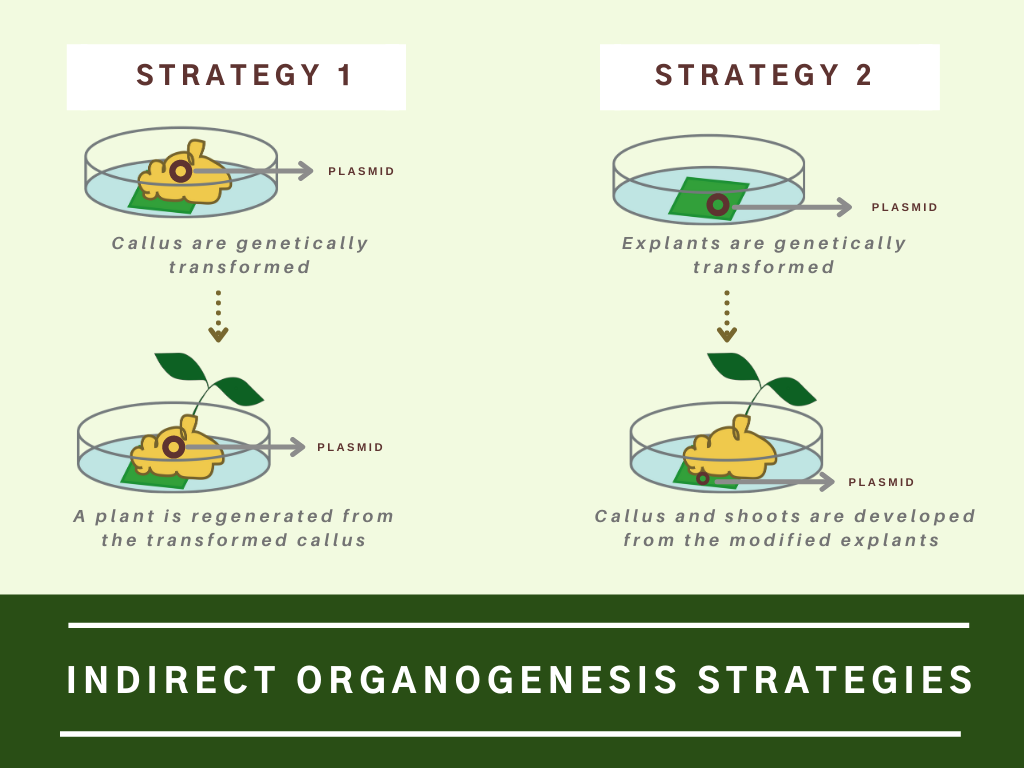
Indirect Organogenesis
The process in which plant organs are derived from a calli mass (tissue formation occurring on a plant wound or at the site of a cut) in the explant is termed indirect organogenesis.
Indirect organogenesis has been used to produce transgenic plants in two ways:
- A plant can be regenerated from a transformed callus.
- The initial explant is transformed, and the callus and shoots are developed from the modified explants.
Typically, indirect organogenesis is more critical for transgenic plant production.
Direct Organogenesis
The production of direct buds or shoots from tissue with no intervening callus stage is called direct organogenesis.
Direct organogenesis has been used to propagate plants with improved multiplication rates (high number of plant per explant) thus reducing operational costs. It has also been used to produce more transgenic plants. And, most importantly, it has been used for clonal propagation (genetic multiplication of a cultivar without sexual reproduction) since it ensures the production of uniform planting material without genetic variation.
Direct Somatic Embryogenesis
In direct somatic embryogenesis, embryos are formed directly from a cell or small group of cells without an intervening callus production. However, direct somatic embryogenesis is generally a rare event in tissue culture. The biological mechanism of how direct SE occurs is not clear and it happens at random for some tested plants.
Indirect Somatic Embryogenesis
Indirect somatic embryogenesis is a process where a callus is first produced from the explant, and then embryos are formed from the callus tissue or a cell suspension culture. This technique is commonly used in transgenic experiments.
Phases of organogenesis and somatic embryogenesis
Organogenesis
Organogenesis can be split into three phases:
- In the first phase, cells in the explants acquire 'competence' which is defined as the ability to respond to hormonal signals of organ induction. During this phase, the process leading to organogenic competence is called dedifferentiation where differentiated cells become undifferentiated.
- During the second phase, competent cells in cultured explants are destined and determined for specific organ formation under the influence of the phytohormones balance.
- Lastly, the third phase is where morphogenesis proceeds independently of the exogenously supplied phytohormones. It means, plant organs take the shape of roots or shoots when phytohormones are removed from the culture med
Somatic embryogenesis
SE can be split into four phases:
- The first phase is where embryogenic masses (callus retaining cells with embryogenic competence) initiate from vegetative cells or tissues.
- In the second phase, embryogenic cell lines (a group of cells with an embryogenic fate) are maintained and developed.
- The third phase involves somatic embryo formation (embryo undergoes globular, heart shaped, torpedo and cotyledonary stages), and maturation (accumulation of reserve substances).
- Somatic embryos are converted (germinated) into viable plantlets.

Advantages and disadvantages of organogenesis and somatic embryogenesis
|
Organogenesis |
Somatic embryogenesis |
|
Advantages |
plants have been propagated using multiple bud induction organogenesis.
|
|
Disadvantages |
|
|
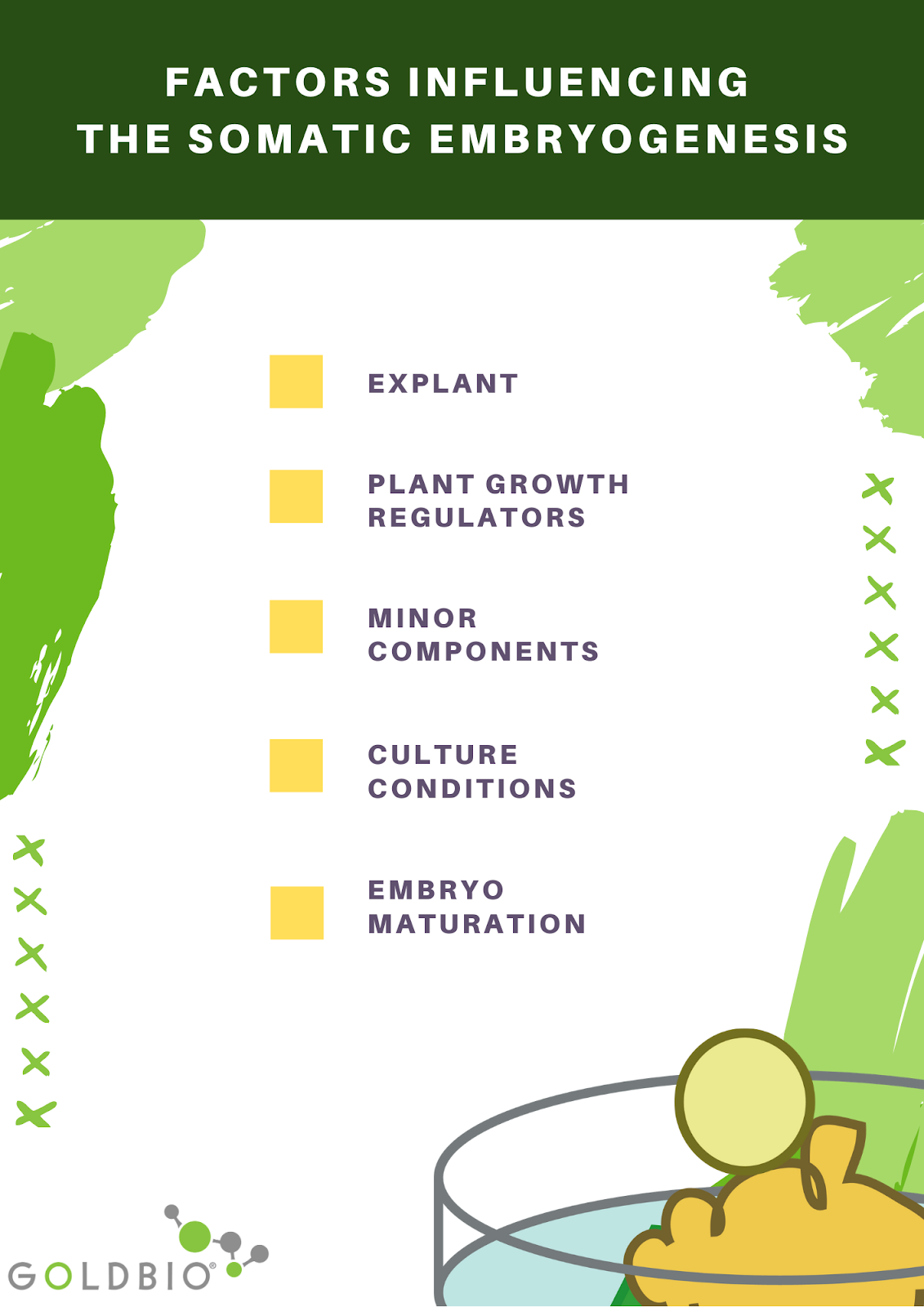
Factors influencing somatic embryogenesis
There are five factors influencing somatic embryogenesis: explant, plant growth regulators, minor components, culture conditions, and embryo maturation.
Explant
The type of explant and the age of the explant can have an impact on the success of somatic embryogenesis.
Young explants especially yield more somatic embryos than older explants.
Another challenge encountered is that, even when you’re choosing different explant tissue (root, flower, shoots, etc.) from the same mother plant, the embryogenic calli are produced at different frequencies.
Here it is necessary to test different concentrations of growth regulators for the induction of somatic embryos.
Furthermore, explants coming from different cultivars of the same species can present varied responses to SE induction. Again, the evaluation of cultivar-specific explants in similar culture conditions is required to determine those with positive responsiveness to somatic embryo production.
Plant growth regulators
If you supplement your media with auxins, it can help promote callus proliferation and inhibit differentiation.
And if you remove or decrease auxins, this will help the development of somatic embryos. Auxins are also responsible for establishing cell polarity in the embryo (apical and basal axis).
Something else we want to point out is that, although cytokinins are also suitable candidates for induction of somatic embryogenesis, fewer cytokinins are used to stimulate SE.
In general, it has been suggested that the cytokinin thidiazuron-TDZ is more effective than other cytokinins for somatic embryogenesis.
Minor components
Amino acids like glutamine, proline, tryptophan, polyamines (putrescine, spermidine), and brassinosteroids (e.g., 24-epibrassinolid) have been reported to enhance somatic embryogenesis in some species.
The potential of these compounds has been attributed to their diverse role in cellular processes such as precursor molecules for certain growth regulators and induce embryogenic pathways through cell reprogramming processes.
Culture conditions
Light condition is a critical aspect when working with SE. For instance, white light enhances growth but at the same time increases the production of phenolic compounds and abscisic acid levels. These substances induce oxidative reactions generating browning of the tissues.
One way to mitigate these light-derived effects is by using activated charcoal in the medium. For other plant cultures like larches, darkness has proved to be as efficient for SE as with light (Lelu-Walter and Paques, 2009). Under this scenario, light conditions should be tested for SE, mainly for the maturation stage.
Embryo maturation
Maturation is regarded as a crucial stage of embryogenesis. Maturation is a preliminary stage for embryo development, which is essentially required for effective germination.
One critical factor in embryo maturation is water loss. Most of the tissue culture treatments promoting the maturation of the embryo use osmoticums (osmotic stress inducers) like sugars.
For instance, apart from being an energy resource, sucrose reduces the water potential of the culture medium, which ultimately leads to water stress, thereby promoting embryo development during in vitro culture.
In this sense, desiccation treatments (by drying the embryos in a filter paper) are also intended to favor embryo maturation.
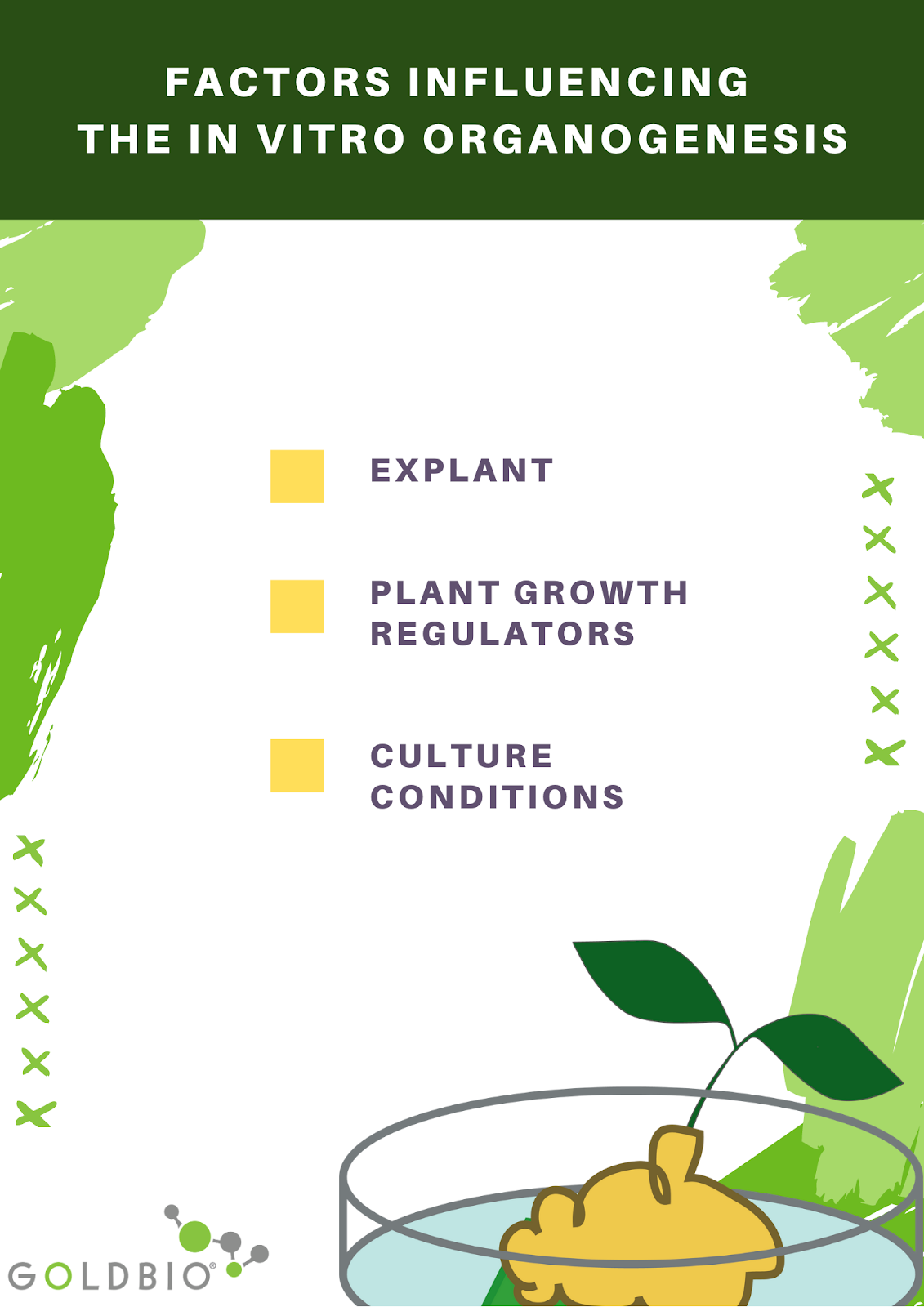
Factors influencing the in vitro organogenesis
There are three primary factors influencing in vitro organogenesis. Among these factors, organogenesis depends mainly on the balance of auxins and cytokinins and the tissue's ability to respond to phytohormones during specific culture conditions.
Explant
A broad range of explants has been used to induce organogenesis. They include leaf disks, petioles, stems, peduncles, stipules, roots, embryos, sepals, and protoplasts. The optimal explant in organogenesis is usually adjusted for each plant species. The next table shows some of the appropriate explants to induce organogenesis in different plant species.
|
Process |
Plant species |
Best explant |
Reference |
Organogenesis |
Strawberry |
Leaf disks |
Husaini et al., 2011 |
|
Chrysanthemum |
Petioles |
Kazeroonian et al., 2018 |
Plant growth regulators
The kind of plant growth regulators (PGRs) and the amount used for plant regeneration varies considerably. In general, a combination of auxin and cytokinin is necessary for successful regeneration.
In recent years, the potent cytokinin TDZ has been commonly used as a unique PGR in the medium favoring the shoot regeneration.
TDZ, a substituted phenyl urea (N-phenyl-1, 2, 3-thidiazol-5-ylurea), exhibits strong cytokinin-like activity and has been shown to be an efficacious regulator of in vitro morphogenesis of many dicot plant species (Ahmed et al. 2012).
Culture conditions
Light is essential in influencing organogenesis, mainly for shoot regeneration. In exceptional cases, darkness favors in vitro organogenesis, as is the case of strawberry shoots (Husaini et al., 2011).
Biotech applications of organogenesis and somatic embryogenesis
Organogenesis has been used mainly for applications like:
- Plant multiplication: This application is ideal especially for clonal propagation. As there is more chance to induce direct organogenesis, desirable traits can be preserved from the mother plant. Transgenic plants produced from organs induced in vitro also retain a great potential to be faithfully cloned if direct organogenesis is promoted.
- Germplasm preservation: bud or shoots can be stored using gelling agents, to preserve and delay the developmental process for the organs. The organs can be later released from the gelling agents to continue the regeneration process.
SE has been used in applications such as:
- Cell selection: as the origin of the embryos can be unicellular, a cell line can be derived from a single cell.
- Plant multiplication: SE is the preferred choice technique to propagate plants, especially genetically transformed plants (by Agrobacterium and other transformation protocols).
- Somatic hybrid regeneration: Through protoplast fusion, two genetic materials can be joined to produce a plant with better traits. Here, SE is also used to regenerate plants from these hybrid protoplasts.
- Regeneration of homozygous plants: plants can be regenerated from pollen or ovules as explants. It produces homozygous plants (plants derived from germinal lines).
- Germplasm preservation: Somatic embryos can be stored as synthetic seed (using gelling agents), to preserve the embryo and resembling the seed structure (embryo plus nutritional substances).
- Virus elimination: As an embryo can be derived from non vascular tissues (usually virus transport around there) and the origin can be unicellular, plants produced in these conditions can be free from virus.

GoldBio support
At GoldBio, we help you achieve your plant regeneration goals!
Visit us at https://www.goldbio.com/collection/shop-by-category and shop our catalog.
References
Ahmed, M. R., & Anis, M. (2012). Role of TDZ in the quick regeneration of multiple shoots from nodal explant of Vitex trifolia L.--an important medicinal plant. Appl Biochem Biotechnol, 168(5), 957-966. https://doi.org/10.1007/s12010-012-9799-0
Arnold, S., Sabala, I., Bozhkov, P., Dyachok, J., & Filonova, L. (2002). Developmental pathways of somatic embryogenesis. Plant Cell, Tissue and Organ Culture, 69, 233-249.
Bhatia, S., & Bera, T. (2015). Somatic Embryogenesis and Organogenesis. In Modern Applications of Plant Biotechnology in Pharmaceutical Sciences (pp. 209-230). https://doi.org/10.1016/b978-0-12-802221-4.00006-6
Feher, A. (2019). Callus, Dedifferentiation, Totipotency, Somatic Embryogenesis: What These Terms Mean in the Era of Molecular Plant Biology? Front Plant Sci, 10, 536. https://doi.org/10.3389/fpls.2019.00536
Frugis, G. (2019). Plant Development and Organogenesis: From Basic Principles to Applied Research. Plants (Basel), 8(9). https://doi.org/10.3390/plants8090299
Hicks, G. (1994). Shoot induction and organogenesis in vitro: a developmental perspective. In Vitro Cell Dev. Biol., 30, 10-15.
Husaini, A., Mercado, J., Teixeira da Silva, J., & Schaart, J. (2011). Review of Factors Affecting Organogenesis, Somatic Embryogenesis and Agrobacterium tumefaciensMediated Transformation of Strawberry. Genes, Genomes and Genomics, 5, 1-11.
Kazeroonian, R., Mousavi, A., Jari, S. K., & Tohidfar, M. (2018). Factors Influencing in Vitro Organogenesis of Chrysanthemum morifolium cv. 'Resomee Splendid'. Iran J Biotechnol, 16(2), e1454. https://doi.org/10.21859/ijb.1454
Lelu-Walter, M.-A., & Pâques, L. E. (2009). Simplified and improved somatic embryogenesis of hybrid larches (Larix × eurolepis and Larix × marschlinsii). Perspectives for breeding. Annals of Forest Science, 66(1), 104-104. https://doi.org/10.1051/forest/2008079
Liang, H., Xiong, Y., Guo, B., Yan, H., Jian, S., Ren, H., Zhang, X., Li, Y., Zeng, S., Wu, K., Zheng, F., Teixeira da Silva, J. A., Xiong, Y., & Ma, G. (2020). Shoot organogenesis and somatic embryogenesis from leaf and root explants of Scaevola sericea. Sci Rep, 10(1), 11343. https://doi.org/10.1038/s41598-020-68084-1
Mazri, M., Meziani, R., Belkoura, I., Mokhless, B., Nour, S. (2018). A combined pathway of organogenesis and somatic embryogenesis for an efficient large‑scale propagation in date palm (Phoenix dactylifera L.) cv. Mejhoul. 3 Biotech, 5, 1-8.
Sugiyama, M. (1999). Organogenesis in vitro. Current Opinion in Plant Biology, 2, 61-64.




