Introduction into Proteinase K
by Tyasning Kroemer, Ph.D.

by Tyasning Kroemer, Ph.D.
Proteinase K is a serine protease commonly used in molecular biology to digest unwanted proteins or inactivate RNases/DNases for preparations of genomic DNA and high-molecular weight nucleic acids.
In this article, we'll take you through what proteases are and how they work, more detail about proteinase K and how it's used.
You can also visit our proteinase K product page to find more information about common applications and usage tips.
Proteases are important reagents in biotechnology and molecular biology laboratories. The use of proteases is often necessary to inactivate protein contaminants in our prepared samples.
Proteases are enzymes that catalyze the cleavage of a protein into amino acids. These enzymes digest a protein by using an amino residue or an activated water molecule as a nucleophile. A nucleophile is an atom of an amino acid that donates an electron to another molecule, causing changes in another amino acid.

Proteases use two types of mechanisms to break a peptide bond:
1. The first mechanism for peptide bond breaking involves the use of an activated water molecule to perform a nucleophilic attack. Examples for these enzymes are aspartic-, glutamic, and metallo proteases.

2. The second mechanism involves the use of a nucleophilic residue to cleave a peptide bond. Serine proteases use an active serine to perform a nucleophilic attack. Cysteine peptidases use the sulfur in cysteine to perform nucleophilic attack. Threonine proteases use a threonine residue as a nucleophile.
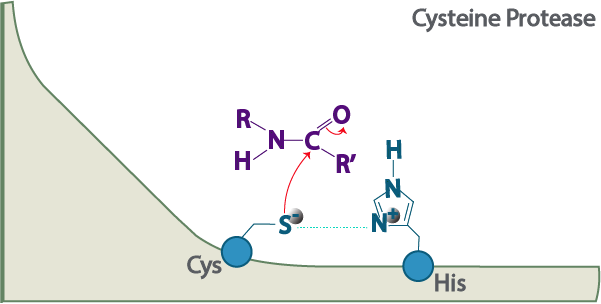
Proteases can be divided into different classes based on the location of their cut, chemical structures, or catalytic mechanism.
Based on the cleavage site, proteases can be classified into:
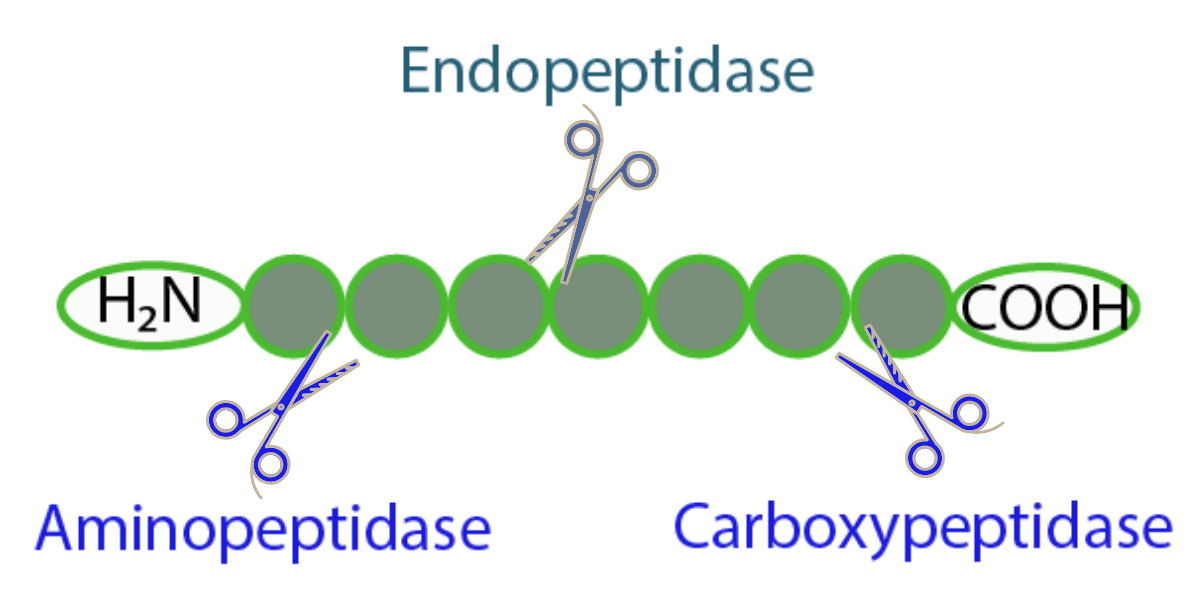
Different Classes of Peptidases. The endopeptidase breaks internal peptide bonds. The aminopeptidase cuts at the amino terminus of a substrate, whereas carboxypeptidase cleaves near a carboxy terminus of a substrate.
However, there is another way to classify proteases. Based on their chemical structures and catalytic mechanisms, there are six classes of proteases:
Serine proteases are proteases characterized by the presence of a reactive serine residue in a catalytic triad. A catalytic triad is a group of three amino acids at the enzyme active site that are involved in catalysis. The catalytic triad allows the transfer of protons into and out of the active site. An example of a catalytic triad for chymotrypsin: Ser 195, His 57, and Asp 102. In this example, the three-letter words represent the amino acid, and the number represents their position.
In these serine proteases, the catalytic triad usually consists of serine, aspartic acid, and histidine amino acids. Some other groups of serine proteases may have a different combination of residues in the catalytic triad, such as Ser-His-Glu, Ser-Lys/His, or His-Ser-His. However, the homology studies and analyses of the active site among serine proteases show that all of these enzymes have a nucleophile serine and the same catalytic mechanism.
The catalytic mechanism of the serine protease chymotrypsin usually represents the catalytic mechanism for all of serine proteases. It is because the structure and mechanism of chymotrypsin is well characterized. Chymotrypsin catalyzes the cleavage of peptide bonds of proteins near the carboxyl group of the aromatic amino acid. The catalytic triad of chymotrypsin consists of Ser 195, His 57, and Asp 102.
The step-wise catalytic mechanism of chymotrypsin:
1. The active Serine (Ser 195) performs the nucleophilic attack on the carbonyl carbon atom of the peptide.

2. The nucleophilic attack causes the formation of tetrahedral intermediate.

3. The tetrahedral intermediate changes into an acyl-enzyme intermediate. The solvent water comes to replace the first amine product.

4. The first amine product leaves and the solvent water replaces it.

5. The reaction produces a second tetrahedral intermediate.

6. The final reaction causes the release of the second amine product and the formation of the active enzyme.

Serine protease consists of large class of proteases based on their structural similarity and functions. Below are some well-characterized groups of serine proteases:
Proteinase K is an endopeptidase belonging to the subtilisin group of serine proteases. In molecular biology laboratories, proteinase K is useful during preparation of DNA or RNA samples by degrading and inactivating proteins.
Proteinase K has five cysteines. Four of the cysteine residues form two disulfide bonds (34-124 and 179-248, respectively) and one lies below one of the catalytic triads. Proteinase K often binds two calcium ions required for full enzymatic activity.
Proteinase K has the catalytic triad consisting of Ser 224, His 69, and Asp 39. The substrate recognition sites are two peptide chains, 99-104 and 132-136. It also has a free Cys 73 near His 69.

Catalytic Triad of Proteinase K from P. album. Catalytic triad model is created with SWISS-MODEL (https://swissmodel.expasy.org).
After the addition of a detergent, all sorts of material is released into lysate including harmful nucleases. To protect DNA from degradation, a protease, in this case proteinase K is used. The reason proteinase K is a good choice is because it’s pretty stable at higher temperatures and it’s broad-spectrum in terms of its targets.
The first step for DNA isolation from mammalian cells and leucocytes is lysing the cells and spinning them down to collect a pellet. Once in suspension, detergent (SDS) is added to disrupt the nuclear membrane. Both histone and non-histone proteins release the DNA after digestion of protein by proteinase K.In addition, proteinase K digests and inactivates ribonuclease in the solution.
After digestion by proteinase K, phenol extractions separate DNA from the lipids and proteins. Hydrophobic lipids stay in the organic phase, whereas the DNA resides in the aqueous phase. Protein (including proteinase K) stays at the organic/aqueous interphase. DNA is extracted from the aqueous layer.Phenol extractions are important to eliminate proteinase K from the high molecular weight DNA.
Proteinase K is useful during DNA isolation from other organisms as well, including:
During RNA isolation, the addition of proteinase K degrades and inactivates even tiny amounts of ribonuclease in the presence of substrate. The additional phenol chloroform and isopropanol precipitation step reduces protein contamination and makes the final pellet easier to re-dissolve.
Proteinase K, RNase/DNase free (Goldbio Catalog #: P-480)
Ahmad, N. N., Cu-Unjieng, A. B., & Donoso, L. A. (1995). Modification of standard proteinase K/phenol method for DNA isolation to improve yield and purity from frozen blood. Journal of Medical Genetics, 32(2), 129. doi:10.1136/jmg.32.2.129
Bertoni, M., Kiefer, F., Biasini, M., Bordoli, L., & Schwede, T. (2017). Modeling protein quaternary structure of homo- and hetero-oligomers beyond binary interactions by homology. Sci Rep, 7(1), 10480. doi:10.1038/s41598-017-09654-8
Biasini, M., Bienert, S., Waterhouse, A., Arnold, K., Studer, G., Schmidt, T., et al. (2014). SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information. Nucleic acids research, 42(Web Server issue), W252-W258. doi:10.1093/nar/gku340
Bienert, S., Waterhouse, A., de Beer, T. A., Tauriello, G., Studer, G., Bordoli, L., & Schwede, T. (2017). The SWISS-MODEL Repository-new features and functionality. Nucleic acids research, 45(D1), D313-d319. doi:10.1093/nar/gkw1132
Blin, N., & Stafford, D. W. (1976). A general method for isolation of high molecular weight DNA from eukaryotes. Nucleic Acids Research, 3(9), 2303-2308. doi:10.1093/nar/3.9.2303
Brown, S. J., & Coleman, M. (2019). Isolation of High Molecular Weight DNA from Insects. In S. J. Brown & M. E. Pfrender (Eds.), Insect Genomics: Methods and Protocols (pp. 27-32). New York, NY: Springer New York.
Ebeling, W., Hennrich, N., Klockow, M., Metz, H., Orth, H. D., & Lang, H. (1974). Proteinase K from Tritirachium album Limber. Eur J Biochem, 47(1), 91-97. doi:10.1111/j.1432-1033.1974.tb03671.x
Gross-Bellard, M., Oudet, P., & Chambon, P. (1973). Isolation of High-Molecular-Weight DNA from Mammalian Cells. European Journal of Biochemistry, 36(1), 32-38. doi:10.1111/j.1432-1033.1973.tb02881.x
Guex, N., Peitsch, M. C., & Schwede, T. (2009). Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: a historical perspective. Electrophoresis, 30 Suppl 1, S162-173. doi:10.1002/elps.200900140
Hedstrom, L. (2002). Serine Protease Mechanism and Specificity. Chemical Reviews, 102(12), 4501-4524. doi:10.1021/cr000033x
Henry, J. M., Raina, A. K., & Ridgway, R. L. (1990). Isolation of high-molecular-weight DNA from insects. Analytical Biochemistry, 185(1), 147-150. doi:https://doi.org/10.1016/0003-2697(90)90270-J
Jackson, D. P., Lewis, F. A., Taylor, G. R., Boylston, A. W., & Quirke, P. (1990). Tissue extraction of DNA and RNA and analysis by the polymerase chain reaction. Journal of Clinical Pathology, 43(6), 499. doi:10.1136/jcp.43.6.499
Jobes, D. V., Hurley, D. L., & Thien, L. B. (1995). Plant DNA isolation: a method to efficiently remove polyphenolics, polysaccharides, and RNA. TAXON, 44(3), 379-386. doi:10.2307/1223408
Kraus, E., & Femfert, U. (1976). Proteinase K from the mold Tritirachium album Limber. Specificity and mode of action. Hoppe Seylers Z Physiol Chem, 357(7), 937-947. doi:10.1515/bchm2.1976.357.2.937
Kraut, J. (1971). 7 Chymotrypsin-Chemical Properties and Catalysis. In P. D. Boyer (Ed.), The Enzymes (Vol. 3, pp. 213-248): Academic Press.
Liles, M. R., Williamson, L. L., Rodbumrer, J., Torsvik, V., Goodman, R. M., & Handelsman, J. (2008). Recovery, Purification, and Cloning of High-Molecular-Weight DNA from Soil Microorganisms. Applied and Environmental Microbiology, 74(10), 3302. doi:10.1128/AEM.02630-07
Lizardi, P. M., & Engelberg, A. (1979). Rapid isolation of RNA using proteinase K and sodium perchlorate. Analytical Biochemistry, 98(1), 116-122. doi:https://doi.org/10.1016/0003-2697(79)90714-0
López-Otín, C., & Bond, J. S. (2008). Proteases: multifunctional enzymes in life and disease. The Journal of biological chemistry, 283(45), 30433-30437. doi:10.1074/jbc.R800035200
McGookin, R. (1984). RNA Extraction by the Proteinase K Method. In J. M. Walker (Ed.), Nucleic Acids (pp. 109-112). Totowa, NJ: Humana Press.
MEROPS - the Peptidase Database. (n.d.). Www.Ebi.Ac.Uk. Retrieved March 27, 2020, from https://www.ebi.ac.uk/merops/cgi-bin/family_index?...
Neitzel, J. J. (2010) Enzyme Catalysis: The Serine Proteases. Nature Education 3(9):21. Retrieved from https://www.nature.com/scitable/topicpage/enzyme-c...
Proteinase K - Worthington Enzyme Manual. (n.d.). Www.Worthington-Biochem.Com. Retrieved March 27, 2020, from http://www.worthington-biochem.com/PROK/default.ht...
Saenger, W. (2013). Chapter 714 - Proteinase K. In N. D. Rawlings & G. Salvesen (Eds.), Handbook of Proteolytic Enzymes (Third Edition) (pp. 3240-3242): Academic Press.
Sweeney, P. J., & Walker, J. M. (1993). Proteinase K (EC 3.4.21.14). In M. M. Burrell (Ed.), Enzymes of Molecular Biology (pp. 305-311). Totowa, NJ: Humana Press.
SWISS-MODEL Repository | P06873. (n.d.). Swissmodel.Expasy.Org. Retrieved March 27, 2020, from https://swissmodel.expasy.org/repository/uniprot/P...
Varma, A., Padh, H., & Shrivastava, N. (2007). Plant genomic DNA isolation: An art or a science. Biotechnology Journal, 2(3), 386-392. doi:10.1002/biot.200600195
Waterhouse, A., Bertoni, M., Bienert, S., Studer, G., Tauriello, G., Gumienny, R., et al. (2018). SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic acids research, 46(W1), W296-w303. doi:10.1093/nar/gky427
Wiegers, U., & Hilz, H. (1971). A new method using ‘proteinase K’ to prevent mRNA degradation during isolation from HeLa cells. Biochemical and Biophysical Research Communications, 44(2), 513-519. doi:https://doi.org/10.1016/0006-291X(71)90632-2
Yang, F., Wang, W., Chen, R. Z., & Xu, X. (1997). A simple and efficient method for purification of prawn baculovirus DNA. Journal of Virological Methods, 67(1), 1-4. doi:https://doi.org/10.1016/S0166-0934(97)00060-8

In the story of cell culture, Mycoplasma is the clear antagonist. These tiny, malleable bacteria lack a rigid cell wall and are notoriously difficult to...

When doing cell culture work, the term Mycoplasma has probably popped up. Mycoplasma is a genus of bacteria that lacks a cell wall and is...

Antibiotics and cell selection agents are used to select for specific cell populations to, for instance, generate a stable cell line or conduct a genetic...

Antibiotics are powerful tools that protect cell culture from contamination and help select for transfected cells. However, antibiotics can also have subtle impacts on resistant...


